Synapses and Dendrites |
|
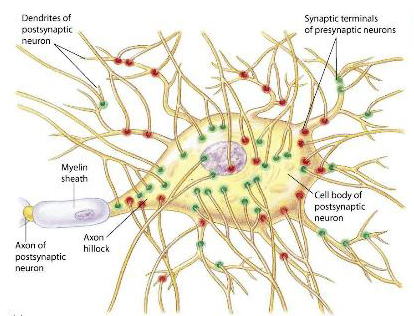
Synapses Synapses are areas of close contact between the axon terminal of one neurone and another cell. Synapses on neurones can occur on the dendrites or the soma (and called axo-dendritic and axo-somatic synapses); most synapses are on the dendrites. The diagram shows synapses on the surface of a motoneurone some of which are red and others green, representing different functions. Excitatory synapses release a neurotransmitter that produces a transient depolarisation of the membrane - an Excitatory Post-Synaptic Potential (EPSP). It is due to current injected into the post-synaptic cell as a result of a neurotransmitter acting on a ligand-gated ion channel in the post-synaptic membrane. There is a synaptic delay - a fraction of a millisecond between the arrival of a nerve inpulse at the pre-synaptic terminal, and the start of the EPSP. The synaptic delay is the time taken for transmitter to be release, diffuse across the synaptic gap, and act on the post-synaptic receptor (the ligand-gated ion channel). This ion channel makes the membrane permeable to a number of different ions - mainly the simultaneous movement of sodium and potassium ions - for a fraction of a millisecond. The electrotonic properties of the membrane cause the EPSP to return to the baseline resting potential within about 12 msec. The equilibrium potential of the EPSP is about +100 to +110 mV. As a result the ion movements during the EPSP depolarise the post-synaptic membrane. Other synapses inhibit the post-synaptic neurone by producing a transient hyperpolarisation called an Inhibitory Post-Synaptic Potential (IPSP). The mechanism is not dissimilar to that described above, except that the neurotransmitter ('inhibitory transmitter') acts on a different ligand-gated channel that opens the channel to potassium and/or chloride ions. The equilibrium potential of the IPSP is between -70 and -90 mV. As a result ion movements hyperpolarise the post-synaptic membrane. Each synapse either depolarises or hyperpolarises the cell membrane and the two type of effects multiplied by the numberous synaptic inputs to the neurone summate algebraically; the resultant changes in the membrane potential at the initial segment of the axon are crucial for the generation of action potentials. |
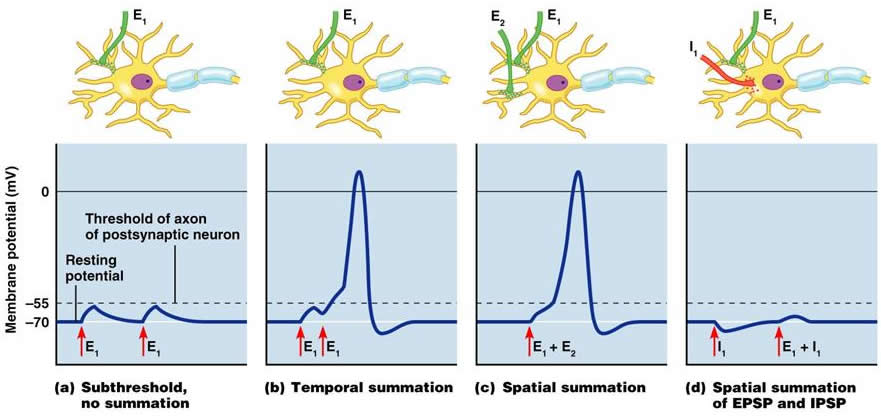
Summation The diagram shows the summation of EPSPs that bring the post-synaptic cell to thershold. It also shows the antagonism between EPSPs and IPSPs that is the essential process underlying the integrative properties of neurones. Axon Hillock The Axon Hillock is the initial segment of the axon - about 25 micrometres in length - and is the site between the perikaryon and the axon at which the cell membrane contains voltage-gated sodium channels. The axon hillock is the site at which action potentials begin and start to pass rapidly down the axon towards its terminal. The axon hillock is therefore the site at which integration of all the EPSPs and IPSPs generated at individual synapses occurs, and the level and duration of depolarisation at the axon hillock determines the frequency of action potentials carried by the axon. |
| Variety of Neuronal Structure | Dendrites on Cerebellar Purkinje Cells |
|
Pre-synaptic endings in the CNS Top |
|
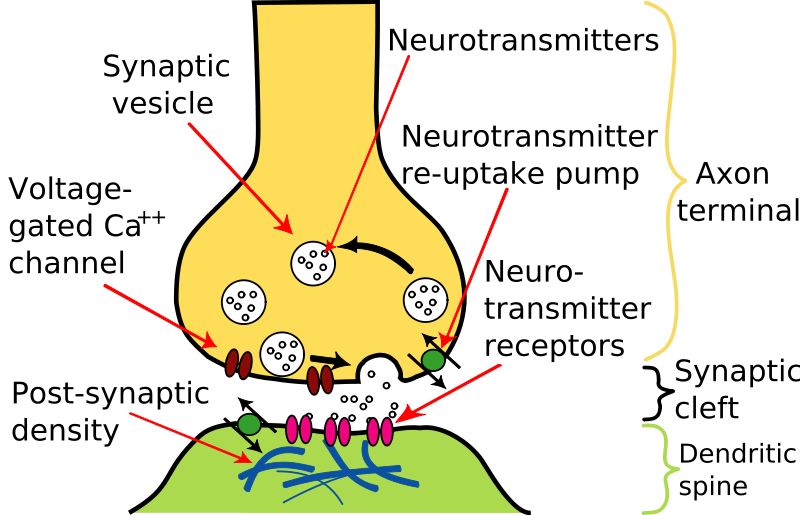
Characteristics of CNS Synapses Synapses typically consist of the end of an axon - the synaptic knob or bouton, a synaptic gap, and the post-synaptic surface on the dendrite or soma of another neurone. The presynaptic terminal contains synaptic vesicles that contain neurotransmitters. Mitochondria are also present in the synaptic bouton as the synthesis of transmitters and the ion pumps require energy. When the action potential arrives at the nerve terminal, voltage gated calcium channels respond to the change in membrane potential and allow calcium to enter the nerve terminal. Synaptic vesicles containing high concentrations of neurotransmitters are found in synaptic boutons, and when calcium enters the bouton, the vesicles fuse with the pre-synaptic membrane and release of the neurotransmitter into the synaptic cleft (or gap). The synaptic gap is generally very narrow, and the post-synaptic membrane is usually thickened by the presence of receptors for the neurotransmitter. So the transmitter diffuses across the narrow gap, which takes a small period of time, the synaptic delay (a fraction of a millisecond). |
|
|
Post-Synaptic Aspects of Neurotransmission in the CNS Top |
|
When the transmitter arrives at the post-synaptic membrane, two events occur: (a) the neurotranmitter is a ligand that binds with specific receptors in the post-synaptic membrane, (b) the neurotransmitter should be removed quickly from the synapse.
|
Neurotransmitters are removed from the synaptic cleft by a variety of different mechanisms in different locations: 1. breakdown, usually by enzymes (as in the Nerve-Muscle Junction), but sometimes by chemical combination with water, as in the case of nitric oxide (NO). 2. by uptake into adjacent cells or the nerve terminal itself, using specialised transporter molecules. Examples include the cell membranes of pre- and post-synaptic cells, and astrocytes. Acetylcholine, an important neurotransmitter in the peripheral nervous system, is only used in a few specific pathways in the CNS. Glutamate is by far the most common excitatory neurotransmitter in the CNS; |
|
Neurotransmitter receptors are proteins that usually consist of several protein subunits. There are many types of neurotransmitter receptor, but there are two main mechanisms by which they achieve their effects: (a) inotropic, in which ions carry electrical current across the membrane by passing through pores that can be open or shut, and (b) metabotropic, in which the neurotransmitter binds to a receptor which in turn activates a cascade of chemical reactions that outlast the presence of the ligand in the synapse. Many metabotropic receptors are coupled to G-proteins, which activate other ion channels or enzymes. |
|
| Neurotransmission using Glutamate in the CNS | Glutamate synthesis from Glutamine Glutamate uptake in presynaptic vesicles Glutamate receptors in the post-synaptic membrane Disposal of Glutamate: the Glutamate-Glutamine Cycle |
|
Inhibitory transmitters produce inhibitory post-synaptic potentials (IPSPs) that hyperpolarise the post-synaptic membrane, and are often released by interneurones that have short axons. Examples (see Chapter 2) are the Ia inhibitory interneurones responsible for reciprocal inhibition in the stretch reflex, the inhibitory interneurones of the Ib clasp-knife reflex, the recurrent inhibition induced by Renshaw cells, and the many types of inhibitory interneurone found in the cerebral and cerebellar cortices. Other types of inhibition include pre-synaptic inhibition, whereby the amount of excitatory transmitter released is reduced as a consequence of activation of an axo-axonic synapse. |
| Two Types of Inhibition | Post-synaptic Inhibition
|
The two main inhibitory transmitters that produce post-synaptic inhibition (IPSPs) in central neurones are GABA and Glycine. GABA (gamma- aminobutyric acid) is the most common inhibitory neurotransmitter, and a derivative of glutamic acid. Glycine, a simple amino acid, is an important inhibitory transmitter in the spinal cord and brainstem. |
| Inhibitory Neurotransmitters : Role of GABA and Glycine in the CNS |
Synthesis of GABA Actions and Receptors of GABA Disposal of GABA Glycine and its receptors |
Acetylcholine is a minor, but important, neurotransmitter in the CNS. It is involved in arousal from sleep. Cholinergic neurones of the basal forebrain project throughout the cortex, and increase the excitability of these neurones when people wake up after sleep. Acetylcholine acts mainly on nicotinic receptors in the CNS, and recent developments in the treatment of Alzheimer's Disease are focused on enhancing the effects of cholinergic nerves. |
Amines are also important transmitters in the CNS and they and ACh will be dealt with in detail in the Chapter on the Reticular formation. About 60 different substances in brain tissue appear to be able to function as neurotransmitters, and the roles of these are gradually being elucidated. These include peptides such as Substance P, Cholecystokinin and the Enkephalins. |
| Synaptic Mechanisms that don't use Synaptic Vesicles | Release and Diffusion of Gases (Nitric Oxide or Carbon Monoxide) in the Nervous System. |